PCR
Introduction
The polymerase chain reaction, or PCR, is a technique for making many copies of a particular region of DNA in vitro. The polymerase chain reaction (PCR) is an effective and efficient way to copy small specific sequences of DNA or RNA and has many applications in biology, medicine, and biotechnology.
PCR depends on a thermostable DNA polymerase, Taq polymerase, and requires the use of pairs of oligonucleotides as initiators or primers of DNA specifically designed for the DNA region of interest. In PCR, the reaction is repeatedly subjected to a cycle of temperature changes that allow the production of many copies of the target region.
In general, the goal of PCR is to produce enough DNA of the target region so that it can be analyzed or used in some other way. For example, PCR-amplified DNA can be sequenced, visualized by gel electrophoresis, or cloned into a plasmid for other experiments.
For this reason, one of the most important parameters for success in PCR amplification is the correct design of the oligonucleotide / primer pair. Because if these are not well designed, the PCR will not work properly.
Therefore, we will finally analyze the main variables to take into account when designing primers/oligonucleotides, among them:
- Melting temperature.
- Oligonucleotide size.
- Specificity.
- Complementarity in the oligonucleotide sequence.
Taq polymerase
Like DNA replication in an organism, PCR requires a DNA polymerase enzyme that produces new DNA strands by using existing strands as a template. The DNA polymerase normally used in PCR is called Taq polymerase, after the heat-tolerant bacterium from which it was isolated (Thermus aquaticus).
T. aquaticus lives in hot springs and hydrothermal vents. Its DNA polymerase is very thermostable and its greatest activity occurs near 70°C (a temperature at which human or E. coli DNA polymerase would not function). Taq polymerase is ideal for PCR thanks to this thermal stability. As we will see, PCR repeatedly uses high temperatures to denature the DNA template, or separate its strands.
PCR primers
Like other DNA polymerases, Taq polymerase can only make DNA if there is a primer, a short sequence of nucleotides that provides a starting point for DNA synthesis. In a PCR reaction, the region of DNA that will be copied, or amplified, is determined by the primers that the researcher chooses.
PCR primers are short pieces of single-stranded DNA, generally about 20 nucleotides long. In each PCR reaction, two primers are used that are designed to flank the target region (the region to be copied). That is, they are given sequences that will make them bind to opposite strands of the DNA template only at the ends of the region to be copied. The primers bind to the template through base complementarity.
When the primers bind to the template, the polymerase extends them and the region between them is copied.

Both primers point “inward” when binding, that is, in the 5’ to 3’ direction toward the region to be copied. Like other DNA polymerases, Taq polymerase can only synthesize DNA in the 5’ to 3’ direction. As the primers are extended, the region between them is copied.
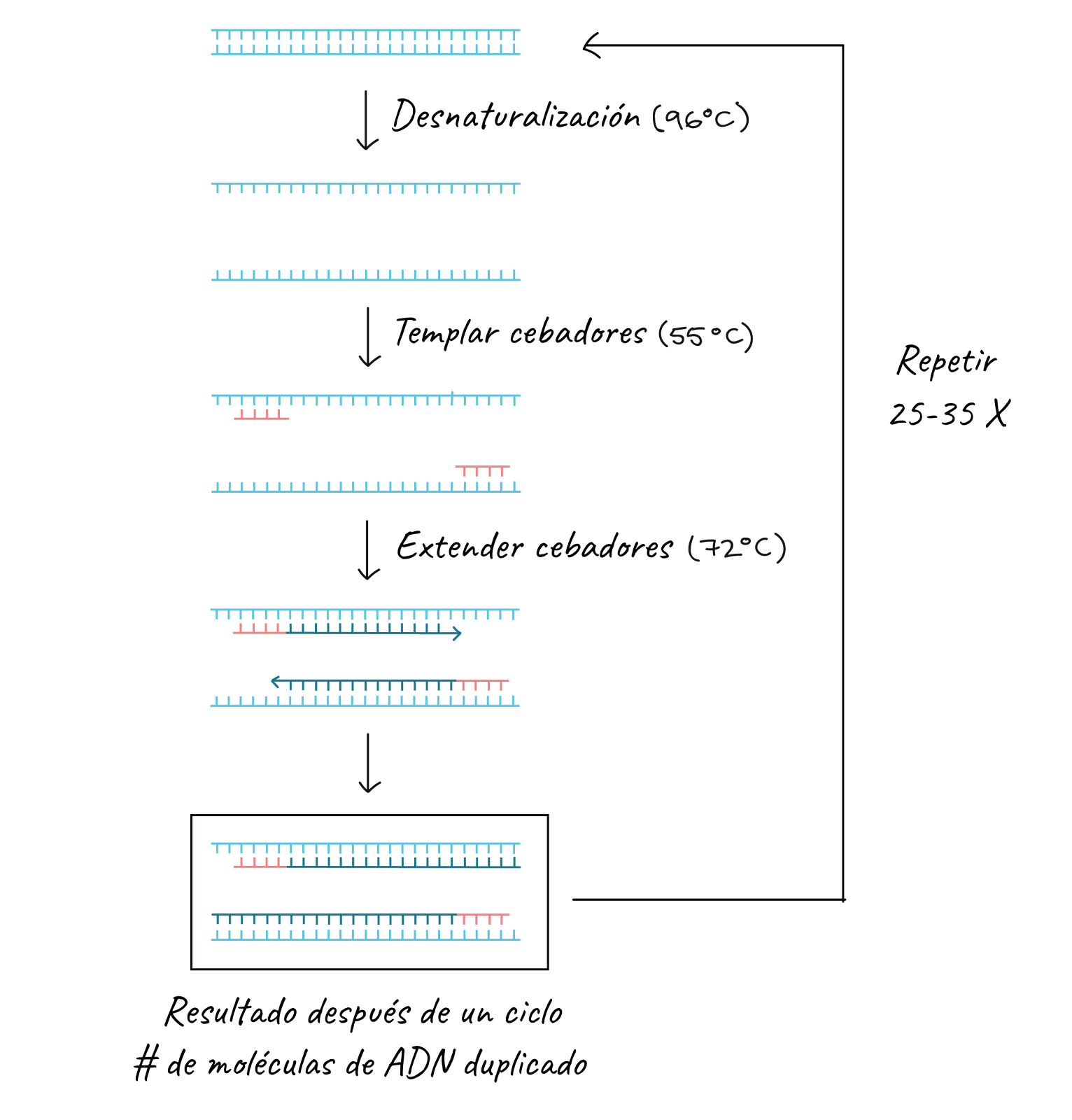
The steps of PCR
As we have already said, the key ingredients for a PCR reaction are Taq polymerase, primers, template DNA, and nucleotides (the building blocks of DNA). The ingredients are placed in a tube, together with the cofactors that the enzyme needs, and are subjected to repeated cycles of heating and cooling that allow DNA synthesis.
The basic steps are:
- Denaturation (96°C): the reaction is heated quite a bit to separate, or denature, the DNA strands. This provides the single-stranded templates for the next step.
- Annealing temperature (55 - 65°C): the reaction is cooled so that the primers can bind to their complementary sequences on the single-stranded DNA template.
- Extension (72°C): the reaction temperature is raised so that Taq polymerase extends the primers and thus synthesizes new DNA strands.
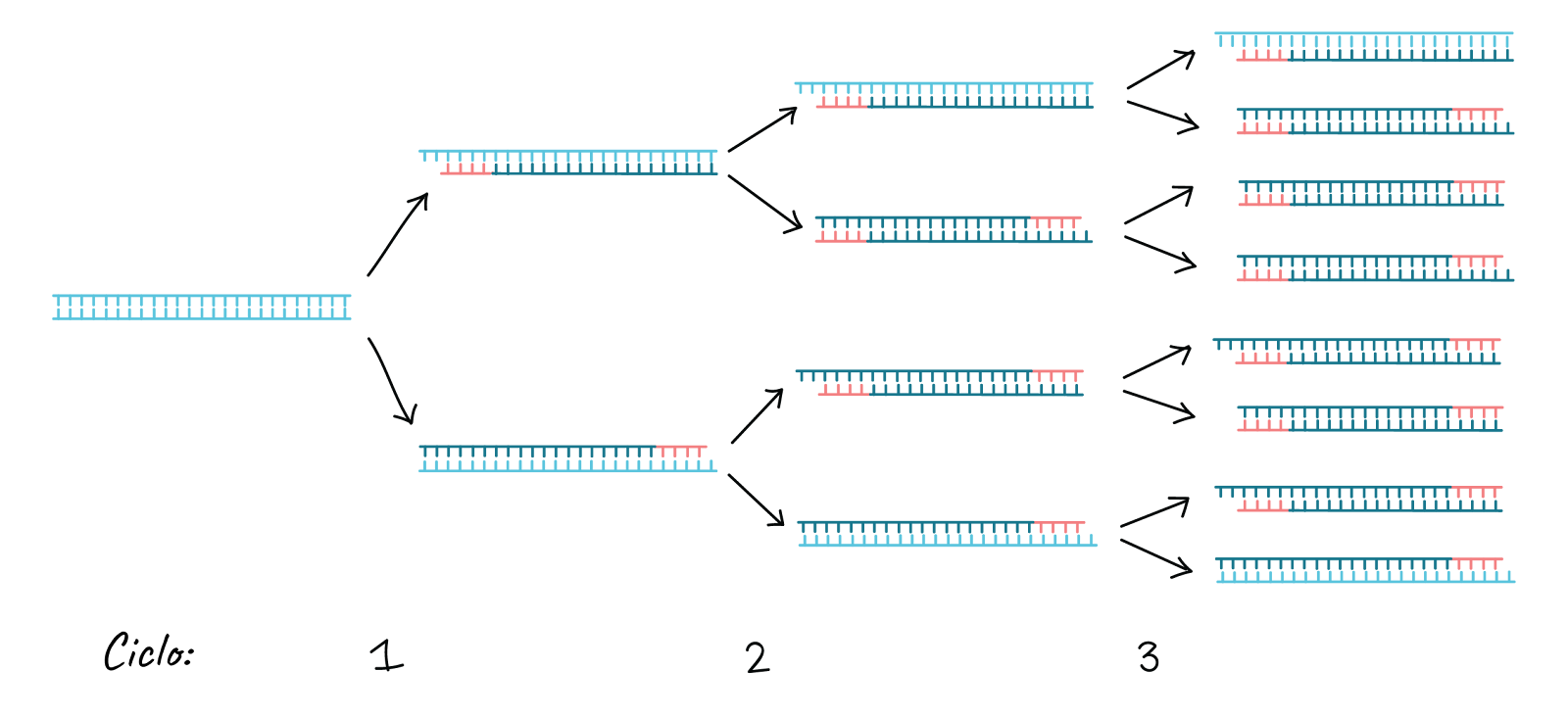
This cycle is repeated 25-35 times in a typical PCR reaction, which generally takes 2-4 hours depending on the length of the DNA region being copied. If the reaction is efficient (works well), it can produce billions of copies from one or a few copies of the target region. This is because not only is the original DNA used as a template in each cycle. In fact, the new DNA produced in one round can serve as a template in the next round of DNA synthesis. There are many copies of the primers and many molecules of Taq polymerase floating in the reaction, so the number of DNA molecules can almost double in each cycle. The following image shows this pattern of exponential growth:
Main variables when performing a PCR
Melting temperature
The melting temperature (Tm) is the temperature at which half of the DNA double strands are denatured. It is important to take into account that in a PCR reaction there are two oligonucleotides and both should be designed so that they have similar Tm. If the oligonucleotides do not have similar Tm, the amplification will be less efficient or may even fail to work, since the oligonucleotide with the higher Tm could anneal non-specifically at low temperatures while the oligonucleotide with the lower Tm may not anneal efficiently at higher temperatures.
The melting temperatures of the oligonucleotides are calculated very accurately with thermodynamic calculations using the following formula:
Tm oligonucleòtids = ΔH [ΔS+ R ln (c/4)] – 273.15°C + 16.6 log 10 [K+]Where H is the enthalpy and S the entropy for the formation of the helix, R is the molar constant and c is the concentration of the oligonucleotide. However, any of the oligonucleotide design programs on the market perform this calculation quickly and easily.
In any case, a good approximation (generally valid for oligonucleotides in the range of 18 – 24 bases) can be calculated with an approximation called the Wallace formula:
Tm oligonucleòtids = 2(A+T) + 4(G+C)Create a table showing Tm values for oligonucleotides of various sizes using the Wallace formula, assuming a G/C content of 50%.
Create a table showing Tm values for oligonucleotides of various sizes using the Wallace formula, assuming a G/C content of 50%.
| Size (bases) | Tm = 2(A+T) + 4(G+C) |
|---|---|
| 4 | 12ºC |
| 8 | |
| 10 | |
| 12 | |
| 14 | |
| 16 | |
| 18 | |
| 20 | |
| 22 | |
| 24 | |
| 26 |
Therefore, as can be observed, the melting temperature of an oligonucleotide depends mainly on its length and on the G+C content. Thus, the base composition of the oligonucleotides should be 45% to 55% in G/C. In addition, the oligonucleotide sequence should be chosen so that it does not contain poly G or poly C zones that can lead to non-specific hybridization. Likewise, poly A and poly T rich zones should also be avoided since these regions can cause premature separation of the strands during the annealing process, which would lead to the breakdown of the template/oligonucleotide complex, reducing amplification efficiency.
Oligonucleotide size
The length of an oligonucleotide sequence influences specificity, the melting temperature, and the time required for hybridization of the oligonucleotide to its complementary sequence, therefore it is decisive for the PCR to work well.
The size of the oligonucleotide is proportional to hybridization efficiency: in general, the longer the oligonucleotide, the more inefficient the hybridization will be. If there are few templates with their hybridized (or annealed) oligonucleotide at each step of the PCR, few amplified products will be generated.
The association of nucleic acid strands longer than 16bp is an extremely specific process. If we take into account the combinatorial possibilities of various “probe sequences” given a DNA strand:
| Probe | Probability | | A, C, G or T/U | 1/4 | | Dinucleotide (e.g. AG) | 1/16 | | Tetranucleotide (e.g. AGCT) | 1/256 |
Thus, the probability of finding a 20bp probe is 1/1,099,511,627,776.
So, oligonucleotides usually have a length of 18 to 24 bases. This size makes the oligonucleotides very sequence-specific, provided that the hybridization temperature is optimal.
Specificity
The specificity of the oligonucleotide depends partly on the size of the oligonucleotide. It is evident that a 24-base oligonucleotide is more specific than a 10-base one. Nevertheless, we must choose oligonucleotides that have a unique sequence in the DNA template that we want to amplify. For example: a pair of oligonucleotides designed with a highly repetitive sequence will give rise to a multitude of PCR products of different sizes when the genomic DNA of a species is used as the template DNA. However, these same oligonucleotides can give rise to a single band if we amplify a clone from a DNA library.
Since the Taq polymerase used in PCR is active over a wide range of temperatures, simply having a small homology at the 3’ end of the oligonucleotide can cause non-specific hybridization of it and the extension of the oligonucleotide to take place at low hybridization temperatures.
Complementarity in the oligonucleotide sequence
It is very important that the oligonucleotides do not have intra-oligonucleotide homology of more than 3 base pairs. If an oligonucleotide has a region of self-homology, partially double-stranded structures can form that will interfere with template hybridization. This type of intra-oligonucleotide homology can lead to the formation of secondary structures. In this way, both the binding between two strands of the same oligonucleotide (self-dimer) and the formation of a hairpin within the same oligonucleotide strand (hairpin) may occur.
Another danger is homology between the two oligonucleotides. Partial homology in the middle regions of two oligonucleotides can interfere with hybridization. If the homology occurs at the 3’ end of each oligonucleotide, oligonucleotide dimers (primer-dimer) will form, which will prevent the PCR from working properly.
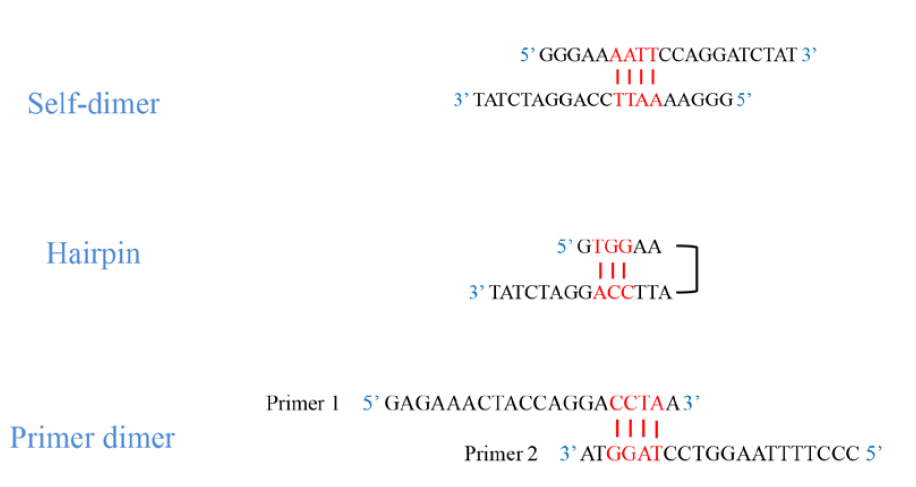
As recommendations, for each of the above homologies we will take into account:
-
The value of the Gibbs free energy (ΔG), which gives an indication of the strength of the secondary structure. It is recommended that the ΔG value be more than –9 for self-dimers and heterodimers.
-
Tm or melting temperature, is the temperature at which an oligonucleotide duplex is 50% in single-stranded form and 50% in double-stranded form. For hairpins, it should be lower than the temperature at which the oligo will be used, and always as low as possible.
-
Another point to consider is where the hairpin occurs, between the ends or in the middle zones. Also whether there are several binding points or few. The best scenario is for them to occur between the middle zones and with few binding points.
Annealing temperature (Ta)
The annealing temperature (Ta) is the estimate of the DNA hybrid and hybrid DNA stability.
As mentioned earlier, the melting temperature of a double strand of nucleic acids is related to its length and G+C content. Thus, the annealing temperature chosen for a PCR depends directly on the length and composition of the primers. It is common to use annealing temperatures 5º below the lowest melting temperature of the pair of primers used (Innis and Gelfand, 1990).
It is important to take into account that a Ta that is too low can lead to non-specificity events of the primers used, such that they end up annealing to sequences different from the one we want to amplify. A Ta that is too high, for its part, can lead to the amplification yield being very low since the probability of annealing is reduced.
Formula to calculate Ta: Ta = 0.3 x Tm(primer) + 0.7 Tm (product) – 14.9 where, Tm(primer) = Melting temperature of the primers and Tm(product) = Melting temperature of the product.
In general, an annealing temperature (Ta) 10º to 15ºC lower than the Tm is usually used.
Practical part
There are a wide variety of programs, many of them free and easily accessible through a web interface. Some of these tools are:
- Oligo (Registration required!!)
- Oligo Calc
- Primer designing tool – NCBI
- Netprimer
- Primer3
- eprimer3
- mfeprimer
During the development of the first part of this practical, we will use the Primer3 tool to design oligonucleotides in silico. Primer3 is an application that is available for free use on different web servers in the world. We will use the implementation of this program offered by University of Tartu & Estonian Biocentre.
The Primer3 browser window has several sections that allow control of a wide variety of parameters (more than 80), related to oligonucleotide design (most of those parameters are rarely modified).
Paste this sequence into the text field at the top of the Primer3 page. Show and explain the results.
GTTCCGGAATTCGCGCGCGTCGACCCACGCGTCCGTCCGCCTCCACCTACCAACCACCATGGCCACGATCGCTATCATCTACTACTCCACCTACGGCCACATCGCCCAACTGGCCGAGGCCGCCAAGAAGGGTGTCGAATCCGTCGACGGCGTGACCGCCGAGATCTACCAAGTGCAGGAGACTCTCTCCGACGAGATTCTGGGCAAGATGCACGCCGCCCCCAAGAAGGACCACCCCATCGCCACTCTTGACACCCTGAAGGAGGCCGATGGCATCCTGTTCGGTTTCCCCACTCGTTTCGGCTCGCTGCCAGCGCAGGTTAAGGCGTTCTTTGACTCGGCCGGTGGCCTGTGGGCTGCTGGGGCTCTGGTAGGCAAGCCTGCCGGTATCTTCTTCAGCACGGGCACTCAGGGCGGCGGTCAGGAGACAACGGCCTTCACTGCCTTGACGTTCCTAGCTCACCAGGGTCTCACCTTCGTGCCACTGGGCTACCGTGCCCCTGAGCTTTTCAACATGGACGAGCTTCACGGTGGATCCCCGTGGGGAGCGGGAACTCTGGCCGGCGGCGATGGCTCTCGCCAGCCATCGAAGCTGGAGCTGACGGTCGCTACCACCCAGGGTAAGTCGTTCGCTAAAGTGGCCAAGAAGCTGGAGGCTTAAGCACTGCGATTATTTGTGTGTGTGTGTGTCTTGGCTCAAGCTAAATTCTAATGCAATAAAATTTAGTTTTACACGGEstás leyendo una vista previa.
Inicia sesión para leer el artículo completo. Cualquier cuenta abre 4 artículos gratuitos al mes; el alumnado y el profesorado leen las páginas de su curso sin límite.
Iniciar sesión